阿伊努人种
波状黑发,体毛多,眼浅褐色(有少数为浅绿色),无蒙古褶,宽脸,鼻宽扁,两眼间距宽。A,B,AB型频率高,O型少,无A2型。Rh 阴性仅4%,NS(MNSS血型系统)出现率为全世界最高者。阿伊努人的起源说法不一,其体质特征及地理分布一直是体质人类学中的谜。
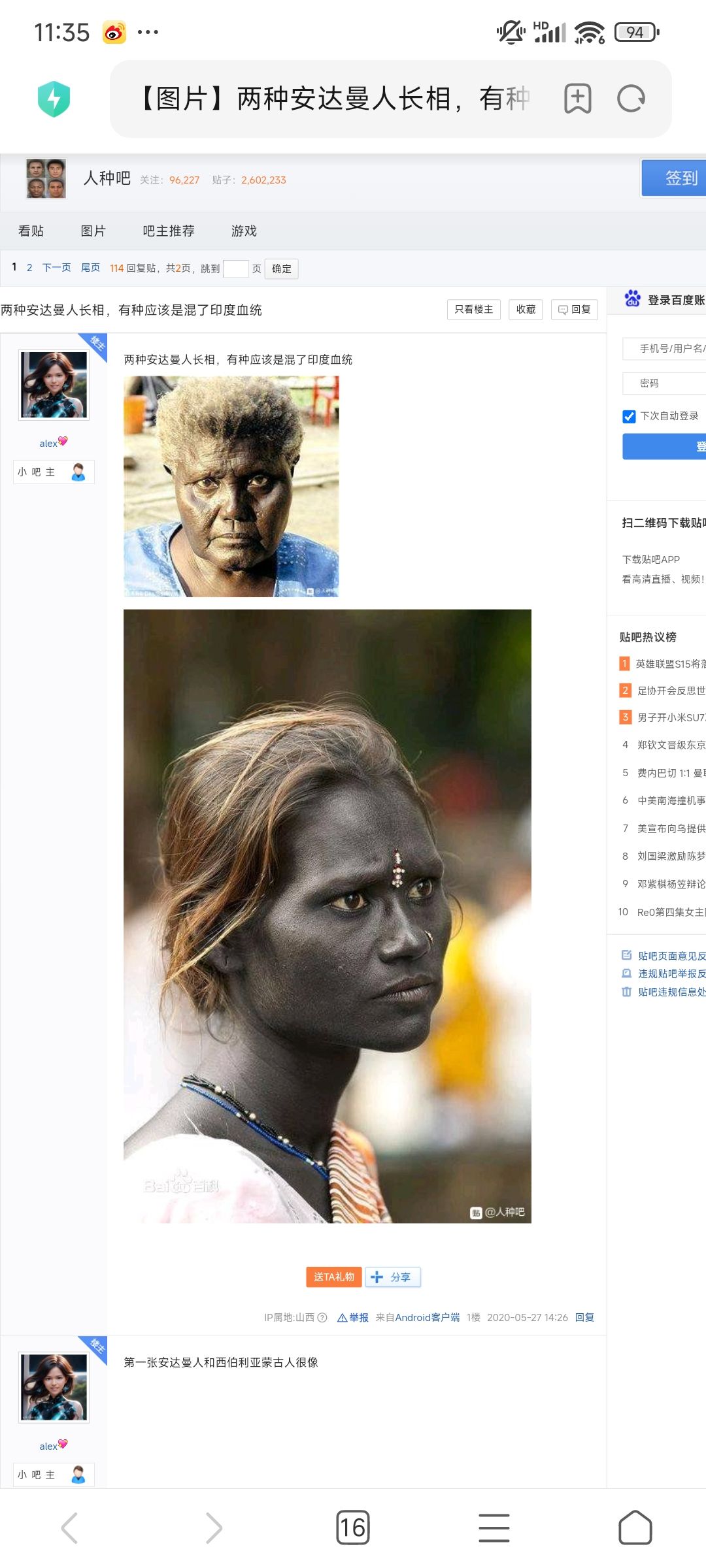
安达曼人种
网上资料少,我只知道该人种A,B,AB型频率高,同时O型很少,而且与阿伊努同为D-Z3660下游,虽然分开很久了。长相的话网传两种长相。上面那种黑鬈发,下面那种波红发,不知是真是假
美拉尼西亚人种
包括斐济、新赫布里底群岛、所罗门群岛、俾斯麦群岛及圣克鲁斯及新几内亚等。过去曾划归澳大利亚人种。美拉尼西亚人种肤色浓黑、波发至鬈发。下颌粗壮、臼齿硕大、容貌粗犷。铲形门齿出现率高,接近蒙古人类。美拉尼西亚人种生活于新几内亚岛北部沿海一带,与内地的巴布亚人(澳大利亚地理人种中的一个地域人种)相比,除肤色更黑之外,还不具鹰钩状鼻子和不像尼格罗人。B型(ABO血型系统)和S型(MNSS血型系统)为主。有地中海贫血基因和6 磷酸葡萄糖脱氢酶缺乏症。β 氢基异丁酸排泄出现率极高(约85%)密克罗尼西亚人种
世界九大地理人种之一。居住于西南太平洋密克罗尼西亚群岛上的居群集团,包括马里亚纳群岛、加罗林群岛、马绍尔群岛、吉尔伯特群岛及瑙鲁岛。过去曾归于澳大利亚人种,或曾认为是澳大利亚土著与蒙古人种(印度尼西亚)的混血后裔。密克罗尼西亚人种肤色中等至浓黑,波型发至鬈曲发(尤其老年),身材较矮小(近似尼格利陀地域人种)。血型特征为B型血(ABO血型系统)出现率高,A型亦常见,N型血(MNSs血型系统)出现率高,M型稍低于N型,Duffy 阳性和Diego 阴性血型常见
波利尼西亚人种
波利尼西亚人和大洋洲人种根本无法等同起来。并非所有的大洋洲人都是棕色人种,例如新西兰的毛利人(属于波利尼西亚人的一支,科学证据表明起源于东亚沿海或者台湾少数民族)属于黄种人。著名的德国人类学家埃克斯坦德特(E.von Eickstedt)提出一种有代表性的的人种分类方法,认为波利尼西亚人属于高加索人种/副人种,是蒙古人种与古代白种人的混合类型,它们的形态特征有些近似于高加索人种,如直发至波发、肤色浅、络缌胡须发育,鼻骨突出,但其面部特征及体毛少等又近似于蒙古人种,如高颅阔颅,面部高阔,高眶型,正颌,从中等身材至高壮身材。国际人类学界所定义的的人种划分法(最新版本),一致认为波利尼西亚人归属蒙古利亚人种。分子人类学证实6000~4000年前,O3a2c*-P164是早期波利尼西亚人最初的父系单倍群,携带东亚母系B4a,发源于东亚沿海迁到台湾,再次渡海经过美拉尼西亚群岛,收留了部分男性居民C2、K3,然后一起迁到太平洋群岛,融合并形成了现代波利尼西亚人。根据血型特征,他们应该是典型的太平洋上的人种。ABO血型系统的B型出现率低(0%~5%),MNSS血型系统N型出现率较高,Duffy阳性血型不甚高,这些都与蒙古人种不同。无ABO系统的A2型和Rh 阴性血型与高加索人种不同,由于与美洲西北海岸印第安人相似,而有波利尼西亚人种起源于印第安人之说。由于地理隔离,人口数目少,其下的地域人种研究不详。但夏威夷岛上的混血人种已构成一个独自的地域人种
澳大利亚地理人种
但其体质特征不同于澳大利亚土著,可能为一地域人种。澳大利亚岛的居民身材多细长、下颌突出、眉嵴显著、牙齿硕大,超过欧洲的尼安德特人化石,体毛多且浓密,头长而窄,下端宽,男子秃顶率高,并伴有小腿无毛现象,与高加索人种近似。儿童的发色有时为浅黄或微红色。ABO血型系统的B型血出现率很低,MNSS血型系统的N型血型出现率极高、无S型血型。与非洲尼格罗人种不同,未发现异常血红蛋白,亦未发现6磷酸葡萄糖脱氢酶缺乏症。在澳大利亚本岛有默伊雷(墨累)地域人种及卡彭(奔)塔里(亚)地域人种生存于内地以游动狩猎为生,他们极为适应夜间的寒冷。N型血型出现率极高(达100%),可能意味着M型血型对澳大利亚人种是不利的。
美洲地理人种
过去曾将其划入蒙古人种,肤色棕褐或棕红,粗直发,发黑眼黑,体毛稀疏,男子秃顶和生痤疮者少,铲形门齿出现率高,颅骨骨缝复杂。并常有缝间骨增加现象,如印加骨(顶枕间骨)。身材较矮小,躯干相对较长,脊柱不弯或很少弯曲,手、脚均较小。无A2、B型和Rh 阴性血型,N型也很少,Diego 阴性出现率高。
总结:
阿伊努人的B型基因是否来自通古斯或尼夫赫有待商榷,但S型基因只能是与美拉尼西亚人同源。我个人看法,尼夫赫也是阿尔冈昆的一支,应该不是从它起源。
至于A型基因和N型基因,那其实大洋洲任何一种人似乎都有可能。