
阿什肯那兹利未人属于父系遗传的犹太群体。
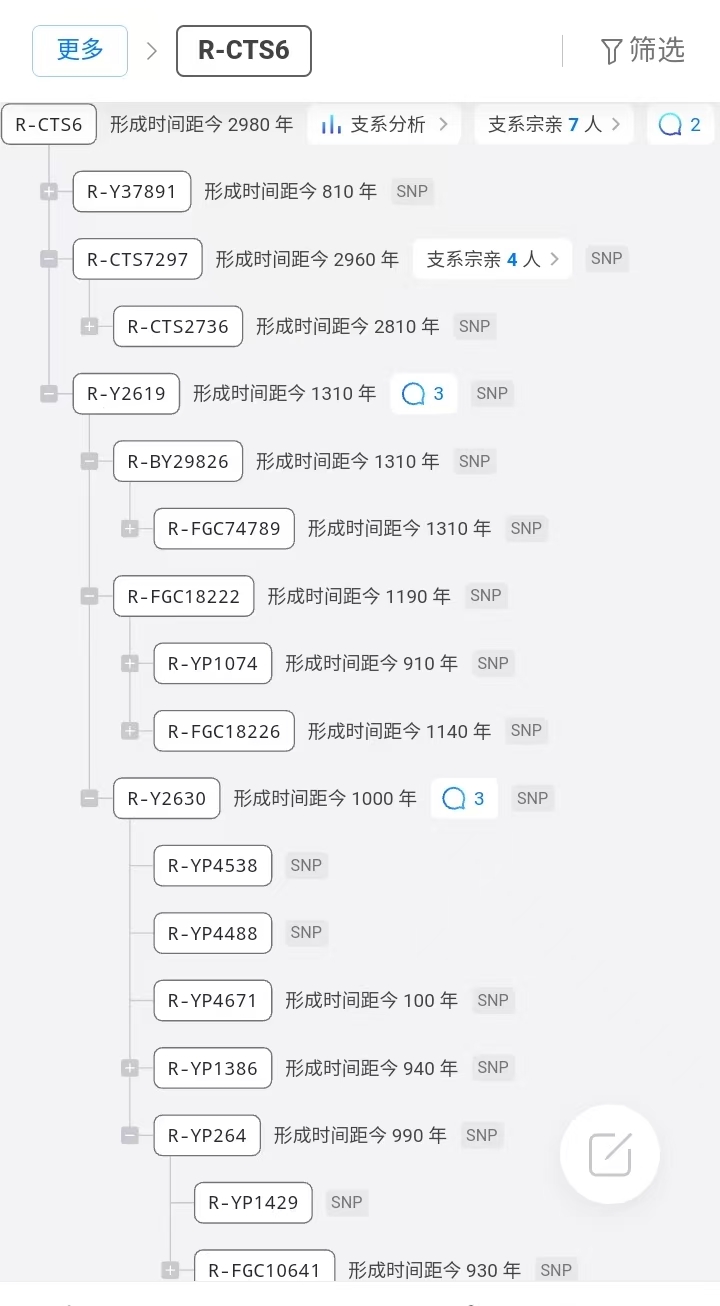
家族名人:以色列政治家本杰明·内塔尼亚胡多次担任以色列总理,根据他兄弟的Ido在FTDNA的DNA测试,属于单倍群R1a-CTS6-Y2619,Y2630犹太分支。
起源研究:早期Y染色体研究显示,阿什肯那兹利未人中有较高频率的非近东单倍群,暗示可能有欧洲男性奠基者事件。但后续全Y染色体序列分析表明,阿什肯那兹利未人Y染色体中的R1a单倍群存在一组独特的核苷酸替换(如M582标记),该标记在东欧样本中几乎不存在,而在近东男性中有一定比例且多样性更高,支持其起源于近东,且可能存在于流散前的希伯来人中。
遗传多样性研究:研究表明,全球约30万自认为是阿什肯那兹利未人的男性中,三分之二可追溯到单个男性祖先。对486个阿什肯那兹和非阿什肯那兹利未人R1a单倍群的Y染色体研究发现,其祖先可能是中东地区自认为是利未人的男性,携带R1a-Y2619谱系,该谱系在希伯来人中可能是一个次要单倍群,约1743年前开始扩张,作为阿什肯那兹人口总体扩张的一部分,随后传播到非阿什肯那兹利未人群体中。
阿什肯纳兹非大祭司的利未人大簇起源猜测:
方案A、与圣经中所提的“赫人”(赫梯人)有关,R1a-Y2619源自印欧语系,近东起源(其上游R-CTS6在东欧非犹太人中极为罕见),融入犹太人的年代非常久远,这与赫梯人的历史比较吻合,在大卫王时代,他们就大量加入希伯来军队,并担任军官,其中有人融入利未人,并随着阿什肯那兹群体的人口增长而成为大族。
方案B、可萨汗国的贵族皈依犹太教,而加入阿什肯那兹利未人中,但,这个推测与其上游CTS6已在近东三千年之久、且在非犹太人中罕见的情况不符。
方案C、在《摩西与一神教》一书中,弗洛伊德通过对摩西传说的深入研究,指出摩西本人并非犹太人,而是一位埃及贵族。这个“埃及”若指喜克索斯人王朝,西亚的赫梯人入侵埃及,建立王朝,就有意思了。摩西并不“口吃”,只是他不精通希伯来语,需要亚伦当翻译,才能跟民众交谈,则这个比科恩家族J1-Y3088更兴旺的利未人家族,疑似摩西本尊!传说犹太人是雅各十二子繁衍的民族,基因检测发现所谓支派是十二个地方行政单位,是一个部落联盟,人员构成十分复杂。类似汉族传说祖先是炎黄二帝、各姓氏是诸侯封国。摩西与亚伦的关系,可能并非亲兄弟,而是结拜兄弟,是盟友。类似姜太公与周文王。
利未人两大家族:摩西家族R-CTS6,Y2619,亚伦科恩家族J-Y3088。
 摘要
摘要阿什肯那兹利未人(Ashkenazi Levites)作为犹太教祭司种姓的重要组成部分,其Y染色体研究为理解犹太人群体的起源、演化和遗传多样性提供了独特视角。本研究基于2013年至2025年间发表的全Y染色体测序数据,深入分析了阿什肯那兹利未人R1a单倍群的分子标记特征、起源假说、遗传多样性模式、地理分布特征以及相关技术争议。研究发现,M582标记作为阿什肯那兹利未人R1a谱系的关键识别标志,在所有采样的阿什肯那兹利未人R1a个体中均有发现,而在922个东欧非犹太男性样本中完全缺失。全Y染色体序列分析揭示了19个独特核苷酸替换定义的阿什肯那兹利未人R1a谱系,其核心为R1a-Y2619分支,共同祖先时间约为1,743年前 。遗传多样性分析表明,全球约30万自认为阿什肯那兹利未人的男性中,约三分之二可追溯到单个男性祖先 。地理分布研究显示,该谱系起源于近东地区,随后作为阿什肯那兹犹太人群体扩张的一部分传播至欧洲,并进一步扩散到非阿什肯那兹利未人群体。然而,研究方法学争议、样本选择偏差以及"犹太基因"概念的社会伦理问题仍需要深入探讨。本研究为阿什肯那兹利未人的遗传历史提供了全面的分子证据,同时也揭示了群体遗传学研究在处理复杂人群历史时面临的挑战。
引言
阿什肯那兹利未人是犹太教中一个特殊的父系遗传群体,根据犹太教传统,他们是利未支派的后裔,负责协助祭司(科恩)进行宗教仪式。在全球约1500万阿什肯那兹犹太人中,约有30万人自认为是利未人,其中约三分之二携带相同的Y染色体谱系 。这一独特的遗传现象引起了人口遗传学家的广泛关注,因为它不仅涉及犹太人群体的起源问题,还关系到我们对人类群体历史、宗教身份与遗传关系的理解。
早期的Y染色体研究显示,阿什肯那兹利未人在东欧最普遍的Y染色体单倍群R1a中表现出显著的奠基者效应。传统观点认为,这一现象支持阿什肯那兹利未人具有东欧起源,可能与中世纪时期犹太人在东欧的大规模定居有关。然而,随着全Y染色体测序技术的发展,研究者们发现了更为复杂的遗传图景。2013年,Rootsi等人通过分析16个完整的R1序列,首次在分子水平上定义了阿什肯那兹R1a谱系的独特性。随后,Behar等人在2017年通过分析486个Y染色体样本,进一步揭示了R1a-Y2619谱系的演化历史和传播路径 。
近年来,随着古代DNA技术的进步和全基因组测序成本的降低,阿什肯那兹利未人的遗传研究进入了新的阶段。2023年,人类Y染色体完整序列的破译更是为这一研究领域提供了前所未有的技术支持 。然而,这些技术进步也带来了新的挑战,包括样本选择的代表性问题、分析方法的标准化问题,以及"犹太基因"概念可能带来的社会伦理争议 。
本研究旨在系统梳理2013年至2025年间阿什肯那兹利未人Y染色体研究的最新进展,深入分析其R1a单倍群的分子特征、起源假说、遗传多样性模式和地理分布特征。同时,我们也将探讨当前研究中存在的方法学争议和伦理问题,以期为该领域的未来发展提供全面的视角。
一、R1a单倍群的分子标记体系与特征
1.1 M582标记的发现与验证
M582标记的发现是阿什肯那兹利未人Y染色体研究的重要里程碑。2013年,Rootsi等人通过对16个完整R1序列的系统分析,识别出了定义阿什肯那兹R1a谱系的19个独特核苷酸替换。其中,M582(也称为CTS2253)被证明是最具诊断价值的标记之一。在对2,834个R1a样本的大规模调查中,研究者发现M582在所有采样的阿什肯那兹利未人R1a个体中均存在,而在922个东欧非犹太男性样本中完全缺失。
这一发现的重要性在于它彻底改变了我们对阿什肯那兹利未人起源的认识。在此之前,研究者普遍认为阿什肯那兹利未人的高频率R1a单倍群支持其东欧起源假说。然而,M582在东欧人群中的缺失,以及它在33.8%的其他R1a阿什肯那兹犹太男性和5.9%的303个R1a近东男性中的存在,强烈暗示了一个完全不同的起源图景。更重要的是,M582在近东男性中表现出显著更高的多样性,这通常被认为是一个谱系起源地的标志。
随后的研究进一步验证了M582标记的可靠性。在对97个随机选择的阿什肯那兹利未人的全Y染色体测序中,研究者发现其中65%携带R1a-M198单倍群,而100%的这些样本都可以被重新分配到R1a-M582细化单倍群。这一结果不仅确认了M582作为阿什肯那兹利未人R1a谱系特异性标记的地位,也为后续的大规模群体研究奠定了基础。
1.2 R1a-Y2619谱系的系统发育特征
在M582标记的基础上,Behar等人在2017年通过分析486个Y染色体样本,进一步识别出了R1a-Y2619谱系作为阿什肯那兹利未人的核心遗传特征 。R1a-Y2619定义了一个包含6个多态位点的分支,其共同祖先时间(TMRCA)估计为1,743年前(95%置信区间:1,334-2,200年前) 。
R1a-Y2619谱系呈现出典型的星状系统发育模式,这与其他阿什肯那兹父系谱系的聚结模式相似 。这种星状模式通常被解释为快速人口扩张的结果,暗示着该谱系可能是阿什肯那兹人群的奠基者谱系之一。值得注意的是,这一扩张模式与利未人的宗教身份没有特殊关联,而是作为阿什肯那兹人口总体扩张的一部分发生的 。
从系统发育的角度来看,R1a-Y2619谱系在R1a-M582内部形成了一个独立的分支。该谱系的所有当代男性携带者,无论他们是否自认为是犹太人、是否为阿什肯那兹人、是否为利未人,都是约1,743年前一位共同男性祖先的直接男性后代。由于来自R1a-Y2619所有分支的当代男性都携带利未人的姓氏,研究者认为这位共同祖先很可能自认为是利未人,并可能携带"Levite"这一姓氏。
1.3 全Y染色体测序技术的应用与进展
全Y染色体测序技术的发展为阿什肯那兹利未人研究带来了革命性的变化。传统的Y染色体研究主要依赖于短串联重复序列(STR)和少量单核苷酸多态性(SNP)标记,这些方法在分辨率和准确性上都有明显的局限。相比之下,全Y染色体测序能够提供数百万碱基对的连续序列信息,使得研究者能够在前所未有的精度上重建Y染色体的演化历史。
在阿什肯那兹利未人的研究中,全Y染色体测序技术的应用主要体现在以下几个方面:首先,它使得研究者能够识别出19个独特的核苷酸替换来定义阿什肯那兹R1a谱系;其次,它提供了足够的序列信息来精确估计共同祖先的时间,如R1a-Y2619的1,743年前的估计 ;第三,它揭示了该谱系内部的精细结构,包括多个亚分支的存在和演化关系。
2023年8月,国际科学家团队完成了人类Y染色体的首个完整无间隙序列,这一成就被发表在《Nature》杂志上 。这项工作填补了Y染色体研究的最后空白,特别是解决了Y染色体上高度重复区域的测序难题。对于阿什肯那兹利未人的研究而言,这一技术突破意味着未来的研究将能够获得更加完整和准确的Y染色体序列信息,从而进一步细化我们对R1a-Y2619谱系演化历史的理解。
1.4 相关标记的变异模式分析
除了M582和Y2619之外,研究者还识别出了多个与阿什肯那兹利未人R1a谱系相关的标记。根据系统发育分析,R1a-Z93(亚洲分支)包含四个主要的子单倍群:M582、M560、Z2125和M780/L657 。这四个子单倍群构成了一个多分叉结构,即使使用10Mb的测序数据也无法完全解析,表明这可能是当前技术条件下无法进一步分辨的系统发育节点 。
在阿什肯那兹利未人的背景下,这些标记呈现出独特的变异模式。例如,DYS456位点在区分阿什肯那兹利未人群体和近东单倍型组方面具有显著的鉴别力。在R1a-M582单倍群内部,只有一个来自阿什肯那兹犹太R1a-M582全基因组样本的STR图谱在DYS456位点显示15个重复,而不是通常的14个重复,这个单倍型距离利未人模态单倍型有两个突变步骤。
此外,研究者还发现了一些与特定历史事件相关的标记。例如,R1a-YP268节点被认为可能代表了Horowitz家族谱系命名创始人的实际重建全Y序列。通过对Horowitz利未人样本的分析,研究者发现三个有详细家谱记录的样本在约402年前聚结,而包括另外两个样本的整个R1a-YP268分支则在约690年前聚结,这与家谱记录的546年前的聚结时间高度吻合。
二、阿什肯那兹利未人R1a单倍群的起源假说
2.1 从东欧起源到近东起源的理论转变
阿什肯那兹利未人起源假说的演变反映了遗传证据与历史叙事之间复杂的互动关系。早期的研究,特别是2003年Behar等人的工作,基于R1a单倍群在东欧的高频率分布,提出了阿什肯那兹利未人具有东欧起源的假说 。这一假说得到了当时遗传数据的支持:阿什肯那兹利未人中有超过50%携带R1a1单倍群,而该单倍群在中东或塞法迪犹太人中并不常见,但在东欧占主导地位。
2003年的研究还发现,阿什肯那兹利未人的R1a1单倍型在微卫星水平上形成了极其紧密的簇,推断的共同祖先在过去2000年内 。与其他犹太群体和非犹太群体的比较表明,这很可能是一个奠基事件,涉及一个或极少数欧洲男性,发生在阿什肯那兹社区初始形成和定居的时期 。这一结论与当时流行的历史观点一致,即阿什肯那兹犹太人在中世纪早期起源于莱茵兰地区,随后向东扩散到斯拉夫土地。
然而,2013年Rootsi等人通过全Y染色体测序技术获得的新证据彻底改变了这一图景。他们发现,M582标记在所有采样的阿什肯那兹利未人R1a个体中都存在,但在922个东欧非犹太男性中完全缺失。这一发现直接挑战了东欧起源假说,因为如果阿什肯那兹利未人的R1a谱系确实来自东欧,那么M582标记应该在东欧人群中也有一定的频率。
更重要的是,M582在5.9%的303个R1a近东男性中存在,并且在近东人群中表现出显著更高的多样性。遗传多样性通常被认为是一个谱系起源地的标志,因为一个谱系在其起源地通常会有更长的演化历史,从而积累更多的变异。这一发现强烈暗示阿什肯那兹利未人的R1a谱系可能起源于近东地区,而不是东欧。
2.2 近东起源假说的分子证据
近东起源假说得到了多方面分子证据的支持。首先,从系统发育的角度来看,R1a-Y2619谱系被嵌入在大量系统发育上接近的中东姐妹分支中,这些分支在伊朗阿塞拜疆人、克尔曼人、雅兹迪人和一个伊比利亚人中都有发现。这种分布模式表明,R1a-Y2619的祖先谱系很可能起源于中东地区。
其次,时间估计也支持近东起源假说。R1a-Y2619的共同祖先时间约为1,743年前,这一时间点完全在罗马流亡散居时期内。考虑到历史上犹太人在公元70年第二圣殿被毁后开始大规模散居,一个在近东起源、随后在散居过程中传播到欧洲的情景是完全合理的。
第三,与其他阿什肯那兹父系谱系的比较分析也支持这一假说。研究发现,R1a-Y2619的扩张模式与其他几个阿什肯那兹祖先谱系相似,如G-M377(约1,223年前聚结)和Q-M242(约1,672年前聚结)。这些谱系都表现出在阿什肯那兹人群体形成时期的同步扩张,暗示它们可能都来自相似的地理来源。
第四,古代DNA研究提供了额外的支持证据。2023年发表的一项关于古代迦南人的研究显示,现代阿拉伯人和犹太人超过一半的DNA来自迦南人和其他古代近东居民。这一发现表明,现代犹太人群体确实与古代近东人群有着密切的遗传联系,为阿什肯那兹利未人的近东起源提供了更广泛的背景支持。
2.3 古代DNA技术的贡献与局限
古代DNA技术的发展为验证阿什肯那兹利未人起源假说提供了前所未有的机会。2022年,Waldman等人发表了对德国埃尔福特中世纪犹太墓地33个14世纪阿什肯那兹犹太人的全基因组研究。这项研究揭示了几个重要发现:首先,埃尔福特个体在遗传上与现代阿什肯那兹犹太人相似,但在东欧相关血统方面表现出比现代阿什肯那兹犹太人更多的变异性;其次,三分之一的埃尔福特个体携带相同的几乎是阿什肯那兹特异性的线粒体单倍群;第三,八个人携带已知影响现代阿什肯那兹人的致病性变异。
这些发现表明,埃尔福特社区已经经历了影响现代阿什肯那兹犹太人的主要规模缩减,即所谓的"阿什肯那兹瓶颈"。然而,埃尔福特的瓶颈比现代的更严重,暗示中世纪阿什肯那兹犹太人存在亚结构。综合来看,这些结果表明阿什肯那兹奠基事件和主要血统来源的获得早于14世纪。
然而,古代DNA技术在阿什肯那兹利未人研究中也存在明显的局限性。首先,目前已发表的古代DNA样本主要来自欧洲的中世纪遗址,缺乏来自近东地区的古代利未人样本。这使得直接验证近东起源假说变得困难。其次,古代DNA的保存条件往往不理想,特别是在温暖潮湿的地区,这限制了来自近东地区古代样本的可用性。
此外,古代DNA研究还面临着伦理和法律挑战。许多可能含有古代犹太人遗骸的考古遗址位于以色列和巴勒斯坦地区,这些地区的政治敏感性使得相关研究变得复杂。同时,犹太教对于处理死者遗骸有严格的宗教规定,这也限制了古代DNA研究的开展。
2.4 历史语言学与考古学的交叉验证
虽然遗传证据为阿什肯那兹利未人的起源提供了重要线索,但要全面理解这一复杂的历史过程,还需要结合历史语言学和考古学的证据。从历史语言学的角度来看,希伯来语和阿拉米语在阿什肯那兹犹太人中的使用和传承表明他们与近东地区的文化联系。特别是犹太教的宗教文献,包括《希伯来圣经》、《塔木德》等,都是用源自近东的语言写成的,这为文化上的近东起源提供了证据。
考古学证据也提供了一些支持。在以色列和巴勒斯坦地区的考古发掘中,研究者发现了许多古代犹太教堂和定居点的遗迹,这些遗迹的年代可以追溯到罗马时期甚至更早。虽然这些考古发现本身并不能直接证明阿什肯那兹利未人的近东起源,但它们提供了古代犹太人群体在近东地区存在和发展的物质证据。
然而,需要注意的是,历史语言学和考古学证据也有其局限性。语言的使用和文化传统可能会在人群迁移过程中发生变化,而考古遗迹的解释往往存在多种可能性。因此,将这些证据与遗传数据相结合,形成多学科的交叉验证,是获得可靠结论的关键。
最新的研究趋势表明,研究者们越来越重视多学科方法的整合。例如,2024年发表的一项研究通过整合遗传、语言和考古学数据,提出了一个关于阿什肯那兹犹太人起源的综合模型 。该模型认为,东欧阿什肯那兹人的祖先最初是通过南部路线来到欧洲的,主要来自安纳托利亚和南欧,而不是传统认为的德国或可萨利亚起源。这一模型得到了遗传学证据的支持,这些证据显示西欧和东欧阿什肯那兹人之间存在差异 。
三、遗传多样性的量化分析与演化模式
3.1 瓶颈效应与奠基者事件的时间估算
阿什肯那兹犹太人经历了一个被称为"阿什肯那兹瓶颈"的严重人口缩减事件,这一事件对包括利未人在内的整个阿什肯那兹群体的遗传结构产生了深远影响。通过对现代和古代DNA数据的综合分析,研究者们能够精确估算这一人口瓶颈的时间、规模和特征。
根据2022年发表的对德国埃尔福特中世纪阿什肯那兹犹太人的研究,研究者们推断出了瓶颈的关键参数:瓶颈期间的有效群体大小约为1,563个二倍体个体(95%置信区间:1,364-1,751),瓶颈开始于41代前(95%置信区间:39-43代),持续了20代(95%置信区间:15-24代)。假设每代25年,这意味着瓶颈开始于约1,000年前,正好是早期阿什肯那兹社区形成的时期。
这一估算得到了多个独立研究的支持。通过对常染色体DNA的分析,研究者发现阿什肯那兹基因组中相同片段(IBD)共享的快速衰减与严重瓶颈后的大规模扩张模式一致。这种模式表明,阿什肯那兹人口在经历瓶颈后经历了快速增长,从15世纪初的约5万人增长到19世纪初的500万人,这一现象被称为"人口奇迹"。
对于利未人群体而言,瓶颈效应的影响更为显著。根据Y染色体数据,全球约30万自认为阿什肯那兹利未人的男性中,约三分之二可追溯到单个男性祖先 。这一惊人的遗传一致性表明,在瓶颈期间,利未人的有效男性群体大小可能只有几十人甚至更少。考虑到利未人在犹太社会中的特殊地位,他们可能在人口缩减期间保持了相对的内婚制,从而加剧了遗传漂变的效应。
3.2 共同祖先时间(TMRCA)的精确估计
通过全Y染色体测序技术,研究者们能够以前所未有的精度估计阿什肯那兹利未人R1a谱系的最近共同祖先时间(TMRCA)。R1a-Y2619谱系的TMRCA估计为1,743年前(95%置信区间:1,334-2,200年前),这一估计基于486个Y染色体样本的系统发育分析 。
这一时间估计得到了多种方法的验证。例如,通过对两个阿什肯那兹M582全序列的比较,它们在8.97 Mb的Y染色体区域内有21个差异位点,使用每碱基对每年1×10^-9的突变率,估计这两个序列的分化时间为1,200年前(标准误300年)。虽然这一估计比R1a-Y2619的TMRCA年轻,但它提供了R1a-M582谱系年龄的下限。
更古老的时间估计来自于R1a-M582谱系与其他相关谱系的比较。通过比较阿什肯那兹M582谱系与亚述R1a-Z2122(xM582)序列,研究者估计R1a-M582分支的年龄上限为4,000年前(标准误300年)。这一估计虽然宽泛,但表明R1a-M582谱系的历史可以追溯到古代近东时期。
有趣的是,基于STR数据的聚结时间分析提供了与SNP数据一致的结果。阿什肯那兹犹太人群体的聚结时间为2.4±1.0千年,而仅来自阿什肯那兹利未人的聚结时间为2.6±1.2千年,这与非阿什肯那兹犹太人的3.1±1.5千年相似。这些估计之间的一致性增强了我们对TMRCA估计可靠性的信心。
3.3 奠基者效应的规模与影响
阿什肯那兹利未人的奠基者效应是人类群体遗传学中最极端的例子之一。根据最新的估计,R1a-Y2619谱系在阿什肯那兹总人口中占7.9%,相当于约30万男性。这意味着,在全球约400万阿什肯那兹男性中,有相当大的比例是约1,743年前一位共同男性祖先的后代。
奠基者效应的规模可以通过多种方式来量化。首先,从姓氏的角度来看,R1a-Y2619谱系的所有当代男性携带者都携带利未人的姓氏,如Levi、Levy、Levin、Horowitz等。这表明,这位共同祖先很可能自认为是利未人,并将这一身份通过父系传承给了后代。
其次,从遗传多样性的角度来看,阿什肯那兹利未人的R1a谱系表现出极低的多样性。在R1a-M582单倍群内部,74%的阿什肯那兹利未人携带相同的模态单倍型,这在人类群体中是极其罕见的。相比之下,在非犹太的R1a高频率群体中,如波兰人、白俄罗斯人等,相同的模态单倍型频率通常不超过5%。
第三,从突变负荷的角度来看,阿什肯那兹人群体,包括利未人,携带了大量的致病性变异。研究发现,八名埃尔福特的中世纪阿什肯那兹犹太人携带已知影响现代阿什肯那兹人的致病性变异。这些变异的高频率是奠基者效应的典型特征,因为在小群体中,有害突变更容易通过遗传漂变而固定。
3.4 与其他犹太祭司群体的比较分析
将阿什肯那兹利未人与其他犹太祭司群体,特别是科恩(Cohanim,亚伦后代,所罗门王时期圣殿大祭司撒都家族)进行比较,有助于我们更好地理解利未人群体的独特性。科恩是犹太教中的高级祭司,传统上被认为是亚伦(摩西的哥哥)的后代,而利未人则是利未支派的后代,负责协助科恩进行宗教仪式。
从遗传特征来看,科恩和利未人表现出截然不同的模式。科恩群体主要携带J1a-P58,Y3088单倍群,该谱系在阿什肯那兹科恩中的聚结时间约为2,570年前,表明其扩张始于散居前时期。与利未人的R1a-CTS6,Y2619谱系不同,科恩的J1a-P58谱系在阿什肯那兹和非阿什肯那兹科恩中都有发现,显示出更广泛的地理分布。
有趣的是,虽然科恩和利未人都属于犹太教的祭司种姓,但他们的Y染色体谱系表明,R1a-Y2619利未人种姓成员和J1a-P58科恩种姓成员在圣经叙事的时间框架内并不共享共同的男性祖先。这一发现支持了犹太教传统中关于这两个群体起源不同的说法:科恩被认为是亚伦的直系后代,而利未人则是利未支派(包括摩西的后代)的其他成员。
从人口统计学的角度来看,科恩和利未人也表现出不同的扩张模式。科恩群体的扩张似乎与犹太教的宗教结构密切相关,而利未人的R1a-Y2619谱系的扩张则"与利未人身份没有特殊关系,而是作为阿什肯那兹人口总体扩张的一部分" 。这表明,利未人的R1a-Y2619谱系可能是在阿什肯那兹人群体形成过程中通过某种方式获得利未人身份的;也可能是利未支派中,能跟亚伦家族相提并论的摩西家族。
此外,从遗传多样性来看,科恩群体表现出比利未人更高的多样性。虽然科恩也有一个占主导地位的单倍群(J1a-P58,Y3088),但该群体中还存在其他多个单倍群,表明其起源可能更加复杂。相比之下,利未人的R1a-Y2619谱系表现出极端的遗传一致性,这可能反映了不同的历史过程。
四、地理分布与传播路径研究
4.1 现代地理分布格局
阿什肯那兹利未人R1a-Y2619谱系的现代地理分布呈现出明显的欧洲中心化特征。根据最新的统计数据,该谱系在不同地区的分布频率如下:波兰(23.51%)、德国(9.33%)、白俄罗斯(7.46%)、捷克共和国(4.85%)。这些数据反映了中世纪以来阿什肯那兹犹太人在东欧和中欧的主要分布模式。
从更广泛的角度来看,R1a单倍群本身在东欧和中欧占据主导地位:波兰(57.5%)、白俄罗斯(51%)、俄罗斯(46%)、乌克兰(45%)、斯洛伐克(41%)。然而,需要强调的是,虽然R1a单倍群在这些地区普遍存在,但阿什肯那兹利未人的R1a-Y2619亚谱系是独特的,不能简单地将其归因于与当地非犹太人群体的基因交流。
现代阿什肯那兹犹太人的全球分布反映了19世纪末到20世纪的大规模移民潮。目前,除了以色列外,阿什肯那兹犹太人主要分布在:美国(约600万)、法国(20万)、加拿大(24万)、英国(26万)、德国(20万)、俄罗斯(约15万)等国。这些移民模式直接影响了R1a-Y2619谱系的现代地理分布。
值得注意的是,R1a-Y2619谱系的分布并不完全与阿什肯那兹犹太人的整体分布重合。在一些传统的阿什肯那兹犹太人聚居地,如立陶宛、拉脱维亚等波罗的海国家,由于历史上的大屠杀和人口迁移,该谱系的频率可能已经发生了显著变化。因此,现代的地理分布模式可能并不能完全反映历史上的传播路径。
4.2 历史传播路径的重建
基于遗传学证据和历史记录,研究者们重建了阿什肯那兹利未人R1a-Y2619谱系的历史传播路径。根据最新的研究,该谱系的变异产生于约2,900年前,随后在公元700年左右阿什肯那兹人口在法国奠基时进入该群体 。
传播路径的第一阶段是从中东到西欧。研究者认为,R1a-Y2619谱系的共同祖先生活在中东地区,可能是在古代以色列或周边地区。在罗马时期,特别是公元70年第二圣殿被毁后,犹太人群体开始大规模散居。在这一过程中,携带R1a-Y2619谱系的利未人可能通过贸易路线或宗教朝圣活动逐渐向西迁移。
第二阶段是在西欧的定居和扩张。根据历史记录,阿什肯那兹犹太人的历史始于10世纪的莱茵兰(德国西部)和法国北部地区 。在这个时期,阿什肯那兹社区开始形成,R1a-Y2619谱系作为奠基者谱系之一被带入该群体。研究表明,该谱系的携带者"可能是阿什肯那兹父系奠基者之一",其扩张是"作为阿什肯那兹人口总体扩张的一部分" 。
第三阶段是向东欧的迁移。11世纪到13世纪的十字军东征后,阿什肯那兹犹太人开始向东迁移到斯拉夫土地(如波兰、立陶宛、俄罗斯) 。这一迁移过程持续了几个世纪,最终形成了历史上著名的"定居点范围"(Pale of Settlement)。在这个过程中,R1a-Y2619谱系随着阿什肯那兹人群体一起向东扩散。
一个特别有趣的案例是Horowitz家族的迁移路径。根据家谱记录和遗传学分析,Horowitz利未人的祖先在约1400年从西班牙的赫罗纳(Girona)迁移到捷克的霍罗维采(Horovice) 。这一迁移路径反映了中世纪时期犹太人群体的流动性,也为我们理解R1a-Y2619谱系的传播提供了具体的例子。
4.3 全球R1a分布模式与阿什肯那兹谱系的独特性
R1a-M420是人类Y染色体上分布最广泛的单倍群之一,但它在欧洲和亚洲的亚结构一直 poorly characterized。通过对欧亚大陆126个人群16,244名男性的大规模研究,研究者识别出了2,923个R1a-M420 Y染色体,并将其分为两个主要群体:一个主要在欧洲(R1a-Z282),另一个局限于中亚和南亚(R1a-Z93)。
R1a-Z282主要分布在欧洲,特别是东欧和中欧地区。该分支下包括多个子单倍群,如R1a-M458(在捷克、斯洛伐克、波兰超过20%)、R1a-M558(在波兰和白俄罗斯超过26%)等。这些子单倍群的地理分布与历史上斯拉夫民族的扩张密切相关。
R1a-Z93则主要分布在亚洲,包括中亚、南亚和西伯利亚地区。该分支下的重要子单倍群包括R1a-M780(在印度、巴基斯坦、阿富汗超过30%)、R1a-Z2125(在吉尔吉斯斯坦和阿富汗普什图人中超过40%)等。这些分布模式反映了不同的历史迁移事件,包括印欧语系人群的扩张和中亚游牧民族的迁徙。
在这一全球分布格局中,阿什肯那兹利未人的R1a-Y2619谱系表现出显著的独特性。首先,它属于R1a-Z93(亚洲)分支,但却主要分布在欧洲;其次,它在近东地区(可能的起源地)的频率很低(5.9%),而在欧洲的阿什肯那兹人群体中频率很高;第三,它形成了一个独立的、高度分化的谱系,与其他R1a-Z93分支在系统发育上相距甚远。
4.4 向非阿什肯那兹利未人群体的传播
阿什肯那兹利未人R1a-Y2619谱系研究中一个特别有趣的发现是,该谱系不仅存在于阿什肯那兹利未人中,也在非阿什肯那兹利未人群体中有发现。研究表明,该谱系"随后传播到非阿什肯那兹利未人" ,但这种传播的机制和时间仍然是一个未解之谜。
从系统发育的角度来看,非阿什肯那兹利未人的R1a-Y2619样本并不形成独立的聚类,而是分散在阿什肯那兹利未人的系统发育树中。具体来说,五个非阿什肯那兹利未人和一个伊拉克犹太人并没有形成独特的系统发育簇,而是分散在阿什肯那兹利未人样本中。这一模式表明,非阿什肯那兹利未人获得R1a-Y2619谱系的时间相对较晚,可能是通过与阿什肯那兹群体的基因交流实现的。
传播的可能机制包括:第一,宗教皈依或身份认同的改变。在犹太教历史上,存在不同犹太群体之间的交流和融合,一些非阿什肯那兹利未人可能通过某种方式获得了阿什肯那兹利未人的血统;第二,婚姻和基因交流。虽然犹太教传统上强调父系传承,但历史上可能存在阿什肯那兹利未人与非阿什肯那兹犹太人之间的通婚,从而导致该谱系的传播;第三,文化传播。在某些情况下,利未人的身份可能通过文化传承而非严格的父系遗传来传递。
时间估计表明,这种传播可能发生在相对较近的历史时期。由于非阿什肯那兹利未人的R1a-Y2619样本都位于阿什肯那兹利未人系统发育树的内部节点,而不是形成基部分支,这表明传播事件发生在R1a-Y2619谱系已经在阿什肯那兹群体中建立之后,可能是在中世纪或更晚的时期。
这一发现对我们理解犹太教的历史和文化具有重要意义。它表明,即使在强调父系传承的宗教体系中,基因谱系的传播也可能通过多种途径实现。同时,它也提醒我们,不能简单地通过遗传谱系来推断宗教身份的历史,因为两者可能通过复杂的方式相互作用。
五、技术方法与研究争议
5.1 全Y染色体测序技术的突破与应用
2023年8月,人类Y染色体首个完整无间隙序列的发布标志着Y染色体研究进入了一个新时代 。这项由来自约翰霍普金斯大学等机构的100多名专家组成的国际团队完成的工作,解决了Y染色体上长期存在的测序难题,特别是高度重复区域的组装问题。
在阿什肯那兹利未人的研究中,全Y染色体测序技术已经显示出巨大的优势。例如,通过对486个Y染色体的测序,研究者不仅识别出了定义R1a-Y2619谱系的6个关键多态位点,还能够精确估计其共同祖先时间为1,743年前 。更重要的是,全序列数据使得研究者能够重建完整的系统发育树,揭示了该谱系内部的精细结构和演化关系。
新的测序技术还带来了长读长测序的突破。传统的短读长测序技术在处理Y染色体上的重复序列时存在困难,而长读长技术能够跨越这些重复区域,提供更完整的序列信息。2023年发表的一项研究展示了如何使用MinION Oxford Nanopore技术对7个主要人类Y染色体单倍群进行基因组组装,为未来的大规模研究提供了技术框架。
然而,全Y染色体测序技术也面临一些挑战。首先是成本问题,虽然测序成本在不断下降,但全Y染色体测序仍然比传统的SNP分型或STR分析昂贵得多。其次是数据分析的复杂性,Y染色体的高度重复结构和复杂的演化历史使得序列比对和变异识别变得困难。第三是质量控制问题,特别是在处理古代DNA样本时,DNA降解和污染可能影响测序结果的准确性。
5.2 研究方法学的争议与批评
阿什肯那兹利未人Y染色体研究领域存在着多重争议,这些争议不仅涉及技术方法,还触及研究伦理和社会影响等多个层面。
主成分分析(PCA)的可靠性争议是当前最激烈的争论之一。2022年,Elhaik发表了一篇极具争议的文章,声称PCA结果可能是数据的人工产物,容易被操纵以产生期望的结果。他指出,PCA在群体遗传学研究中被过度依赖,可能导致32,000到216,000项遗传研究需要重新评估。这一观点对许多基于PCA的阿什肯那兹起源研究提出了根本性的质疑。
古代DNA研究的方法学问题也受到批评。例如,2020年Elhaik对一项关于可萨人(Khazars)的古代DNA研究提出了严厉批评,指出该研究在样本识别、测序环境选择和分析方法等方面存在严重问题。他认为,该研究的分析产生了荒谬的结果,没有任何声明得到数据的明确支持。虽然这一批评针对的是可萨人研究,但它也反映了古代DNA研究中普遍存在的方法学挑战。
样本代表性问题是另一个重要争议点。批评者指出,许多研究使用的样本可能存在选择偏差,特别是在确定个体的利未人身份时。由于利未人身份主要通过家族传统和自我认同来确定,可能存在错误分类的情况。此外,一些研究可能倾向于选择那些支持特定假说的样本,从而影响结果的客观性。
统计方法的选择也存在争议。不同的研究可能使用不同的突变率、不同的群体遗传学模型,这些选择都会影响最终的时间估计和演化推断。例如,使用谱系突变率还是进化突变率,使用严格的分子钟还是放松的分子钟,都会导致显著不同的结果。
5.3 "犹太基因"概念的社会伦理争议
"犹太基因"这一概念本身就充满争议,它不仅涉及科学问题,还触及种族、宗教和身份认同等敏感的社会议题。
种族化风险是最主要的担忧。2023年,Kohler在一篇评论文章中警告,关于犹太人遗传接近性的科学研究可能"重新种族化思维" 。她指出,虽然这些研究声称是客观的科学探索,但实际上它们往往将宗教和文化身份简化为生物学特征,这可能强化种族本质主义的观念。
报道偏见问题也引起了广泛关注。2023年,van Straten发表文章批评遗传学家在报道阿什肯那兹犹太人研究结果时存在偏见。他指出,某些重要发现,如阿什肯那兹女性80%的欧洲背景,以及西欧和东欧阿什肯那兹人线粒体DNA之间的显著差异,往往没有被充分报道。相反,一些缺乏证据支持的结论,如阿什肯那兹男性与圣经以色列人的遗传联系,却被反复强调。
商业利益的影响是另一个值得关注的问题。直接面向消费者的基因检测公司,如23andMe和AncestryDNA,在其血统报告中声称存在可检测的"犹太基因",这不仅在科学上值得商榷,还可能加剧社会分化 。这些公司通常只提供欧洲或阿什肯那兹犹太血统的报告,排除了非阿什肯那兹犹太血统,从而强化了一种以阿什肯那兹为中心的犹太历史叙事。
伦理困境在研究实践中也经常出现。例如,当研究结果可能挑战宗教信仰或社会认同的时候,研究者面临着艰难的选择。一些批评者认为,"科恩基因"和"利未基因"虽然可以提供某人是科恩或利未人的强有力遗传证据,但它们在犹太教法(halacha)中没有任何意义 。这种科学发现与宗教传统之间的冲突可能导致身份认同危机和社会分裂。
5.4 未来研究方向与技术展望
面对当前的争议和挑战,阿什肯那兹利未人Y染色体研究领域需要在多个方向上寻求突破和改进。
技术创新是推动该领域发展的关键。随着长读长测序技术的成熟和成本的降低,未来的研究将能够获得更加完整和准确的Y染色体序列。特别是对于古代DNA样本,新的富集技术和损伤修复方法可能使得从降解的古代DNA中获得高质量Y染色体序列成为可能。此外,单细胞测序技术的发展也可能为研究Y染色体嵌合现象和体细胞突变提供新的机会。
多学科整合是提高研究可靠性的重要途径。未来的研究应该更加注重整合遗传学、历史学、考古学、语言学等多个学科的证据,形成相互验证的研究体系。特别是古代DNA研究,需要更加严格的实验标准和质量控制,以确保结果的可靠性。
研究伦理的规范化也是当务之急。研究者需要建立更加透明和负责任的研究规范,包括样本选择的标准、结果解释的原则、与研究群体的沟通机制等。特别是在涉及宗教和民族敏感议题时,需要更加谨慎和尊重的态度。
社会影响的评估应该成为研究的重要组成部分。在开展研究之前,需要评估可能的社会影响,并制定相应的应对策略。这包括如何向公众解释研究结果,如何防止研究成果被滥用,如何保护研究参与者的隐私和尊严等。
最后,我们需要认识到,阿什肯那兹利未人的遗传研究不仅是一个科学问题,也是一个涉及人类多样性、文化传承和社会公正的重要议题。只有在科学严谨性和社会责任感之间找到平衡,这一研究领域才能真正为人类知识的进步和社会的和谐发展做出贡献。
结论
通过对2013年至2025年间阿什肯那兹利未人Y染色体研究的系统梳理和深入分析,我们可以得出以下主要结论:
分子标记体系的精确解析。M582标记作为阿什肯那兹利未人R1a谱系的关键识别标志,在所有采样的阿什肯那兹利未人R1a个体中均有发现,而在922个东欧非犹太男性样本中完全缺失,这一发现彻底改变了我们对该群体起源的认识。R1a-Y2619谱系作为核心分支,通过6个多态位点定义,共同祖先时间约为1,743年前,呈现典型的星状扩张模式。全Y染色体测序技术的应用使得我们能够在前所未有的精度上重建该谱系的演化历史。
起源假说的根本转变。从早期的东欧起源假说转向近东起源假说,这一理论转变得到了多方面证据的支持:M582在近东人群中的更高多样性、R1a-Y2619谱系在系统发育上与中东姐妹分支的密切关系、约1,743年前的共同祖先时间(完全在罗马流亡散居时期内)等。古代DNA研究虽然提供了一些支持,但仍需要更多来自近东地区的古代样本以直接验证这一假说。
遗传多样性的极端模式。阿什肯那兹利未人表现出人类群体中最极端的奠基者效应之一:全球约30万自认为阿什肯那兹利未人的男性中,约三分之二可追溯到单个男性祖先。瓶颈效应的时间估计为约1,000年前,有效群体大小约为1,563个二倍体个体。与科恩群体的比较分析显示,两个祭司群体具有不同的遗传起源和扩张模式。
传播路径的复杂历史。R1a-Y2619谱系从中东起源,约2,900年前产生变异,随后在公元700年左右进入阿什肯那兹群体。传播路径包括从中东到西欧、在西欧的定居扩张、以及向东欧的迁移三个主要阶段。该谱系还通过未知机制传播到非阿什肯那兹利未人群体,但系统发育分析表明这种传播发生在相对较晚的时期。
方法学争议与伦理挑战。主成分分析的可靠性、古代DNA研究的方法学标准、样本代表性等技术问题仍存在激烈争论。"犹太基因"概念可能带来的种族化风险、报道偏见、商业利益影响等伦理问题需要引起高度重视。这些争议提醒我们,在追求科学真理的同时,必须承担相应的社会责任。
展望未来,阿什肯那兹利未人Y染色体研究仍面临诸多挑战和机遇。技术创新,特别是长读长测序和古代DNA技术的进步,将为我们提供更精确的分子证据。多学科整合研究将帮助我们构建更加完整和可靠的历史图景。同时,建立更加规范的研究伦理和社会影响评估机制,确保研究的科学性和社会责任的平衡,是该领域可持续发展的关键。
阿什肯那兹利未人的遗传研究不仅深化了我们对人类群体历史和演化的理解,也为探讨宗教身份、文化传承和生物遗传之间的复杂关系提供了独特视角。在这个充满争议和挑战的研究领域,我们需要以更加开放、严谨和负责任的态度,推动科学进步,促进社会和谐,最终服务于人类知识的增长和文明的发展。
彝伦
2025-10-28添加评论
1 赞